

קליפת הראייה
תבנית אנטומיה ריקה
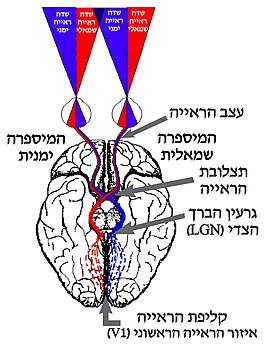
קליפת הראייה (אנגלית: Visual cortex, נקראת גם קליפת המוח הראייתית) היא האזור המרכזי במוח בו מעובד מידע חזותי הנקלט במערכת הראייה. קליפת הראייה מהווה חלק מקליפת המוח, והמוקד שלה הוא באונה העורפית. המידע החזותי שנקלט ברשתית העין מועבר לקליפת הראייה דרך גרעין הברך הצדי שבתלמוס.
קליפת הראייה כוללת בתוכה מספר אזורים, ולכל אזור תפקיד משלו בתהליך עיבוד המידע. אזורי הראייה הראשוניים כמו קליפת הראייה העיקרית מעבדים בעיקר מידע המהווה את הבסיס לראייה, כמו קווים וקצוות, זיהוי תנועה, עומק וצבע, בעוד שאזורי ראייה אחרים בקליפת המוח מעבדים מידע מורכב יותר כמו זיהוי צורות ותבניות, מקומות ופנים.
מבוא
 ערך מורחב – מערכת הראייה
ערך מורחב – מערכת הראייה
קליפת הראייה מקבלת את המידע החזותי דרך מסלול עצבי המגיע מן העין אל אזורי הראייה במוח. המסלול מתחיל למעשה ברשתית, שבה מתרחשת ההתמרה של אור לאותות עצביים. אותות אלה מועברים משתי העיניים אל המוח על ידי עצב הראייה. בתצלובת הראייה, חלק מהאקסונים המרכיבים את עצב הראייה חוצים לצדו השני של המוח, כך שמידע מהצד הימני של שדה הראייה מגיע להמיספרה השמאלית של המוח, ולהפך. מתצלובת הראייה ממשיכים האותות אל גרעין הברך הצדי, שבו מתבצע העיבוד הראשוני של המידע החזותי. צרור של אקסונים, הפרוש לאורך אזור הנקרא מַעֲנִית הַפָּרִישׁ, מוליך את האותות מתאי העצב של גרעין הברך הצדי אל אזורי קליפת הראייה באונה העורפית, שם מתבצע עיבוד מתקדם יותר של המידע החזותי.
נהוג למספר את האזורים השונים המרכיבים את קליפת הראייה בצורה הבאה: V3 ,V2 ,V1 וכו'. אזור V1 נקרא גם "קליפת הראייה העיקרית", והוא תחנת העיבוד הראשונית בקליפת הראייה.
תאי עצב בקליפת הראייה יורים דחפים עצביים כאשר גירוי חזותי מופיע בשדה הקלט שלהם. לכל תא עצב יש כוונון לגירויים בעלי מאפיינים מסוימים, שאליהם הוא מגיב יותר. באופן כללי, בקליפת הראייה העיקרית תאי העצב רגישים למאפיינים פשוטים יותר בשדה הקלט (למשל, פס אור בעל אוריינטציה מסוימת) ובאזורי העיבוד המתקדמים יש כבר כוונון מורכב יותר למאפיינים כמו צבע, צורות גאומטריות ואפילו מקומות ופנים.
קליפת הראייה מקבלת את הספקת הדם שלה בעיקר מהענף הדורבני שבעורק המוח האחורי.
קליפת הראייה העיקרית V1

קליפת הראייה העיקרית V1, הנקראת גם אזור הראייה הראשוני, היא אזור הראייה שנחקר בצורה המקיפה ביותר. אצל כל היונקים אזור זה ממוקם בקוטב האחורי של האונה העורפית, והוא שקול לאזור ברודמן 17. זהו אזור הראייה המוקדם ביותר בתהליך עיבוד המידע החזותי במוח, ומנגנוני עיבוד המידע בו הם הפשוטים ביותר. קליפת הראייה העיקרית מותאמת במיוחד עבור עיבוד מידע חזותי המבדיל עצמים נייחים מעצמים נעים, והיא מצוינת בהכרת דפוסים.
קליפת הראייה העיקרית שקולה בקירוב לאזור האנטומי שמכונה "הקליפה המפוספסת" (Striate cortex). שם זה נגזר מ"הפס של ג'נארי"[1], שהוא פס לבן (הנראה גם לעין בלתי מזוינת) של אקסונים עטופים במיאלין שמגיעים מגרעין הברך הצדי לשכבה מס' 4 של החומר האפור בקליפת הראייה.
גודל הקליפה אצל אדם בוגר הוא 25 סנטימטרים רבועים בקירוב, והמספר הכולל של תאי עצב בקליפה הוערך לסביבות 200 מיליון[2].
מבנה ותפקוד קליפת הראייה העיקרית V1
בקליפת הראייה העיקרית, כמו גם ברשתית עצמה, יש שטח מוקצב של רקמת עצב לכל יחידת שטח בשדה הראייה, אשר מוקדש לעיבוד המידע החזותי מאותו אזור. כך לדוגמה, חלקו העליון של הקליפה בבני אדם (המהווה את הגוש העליון של מענית הפריש) מתאים לחציו התחתון של שדה הראייה, וחלקו התחתון של הקליפה (המהווה את הגוש התחתון של מענית הפריש) מתאים לחציו העליון של שדה הראייה. מיפוי זה של שדה הראייה על פני הרשתית וקליפת הראייה העיקרית (וכך גם באזורי עיבוד ראייתי נוספים) מכונה מיפוי רטינוטופי. זהו מיפוי מאד מדויק - אפילו הכתם העיוור ממופה לקליפת הראייה העיקרית. מבחינה אבולוציונית, התאמה זו היא בסיסית מאד וקיימת ברוב רובם של בעלי החיים להם אזור זה במוח. אצל בני אדם ובעלי חיים שהם בעלי גומה מרכזית ברשתית, חלק גדול מ-V1 מוקדש לחלק קטן במרכז שדה הראייה ("מוקד הראייה"), תופעה הידועה בשם הגדלה קליפתית (Cortical magnification).
לתאי העצב ב-V1 יש את שדה הקלט הקטן ביותר מכל התאים בקליפת הראייה, אולי לצורך קידוד מדויק של מיקום מרחבי.
יכולת הכוונון של תאי העצב ב-V1 לגירוי ספציפי (אליו תא העצב יגיב) משתנה במידה רבה עם הזמן. בהתחלה (החל מ-40 מילישניות מתחילת הצגת הגירוי) לנוירונים העצמאיים ב-V1 יש כוונון חזק לקבוצה מצומצמת של גירויים, כלומר התגובות העצביות יכולות להבחין בשינויים זעירים בכיוון שדה הראייה בתדירות מרחבית (המהירות שבה גירוי משתנה במרחב) ובצבע. זאת ועוד, לתאי העצב ב-V1 בבני אדם ובעלי חיים בעלי ראייה בשתי עיניים יש את היכולת להתכוונן לרק אחת משתי העיניים. ב-V1 ובקליפות חושים עיקריות אחרות, תאי עצב עם יכולות כוונון דומות נוטות לחבור יחד וליצור עמודות קליפה רבות שכבה. דייוויד הובל וטורסטן ויזל היו הראשונים שהציעו את מבנה הקובייה (עליו נרחיב בהמשך) של קליפות שכבה כמודל הארגון עבור שתי יכולות כוונון: שליטה בתזוזת העיניים ואוריינטציה (יכולת כיוון). אולם מודל זה אינו מתחשב בצבע, תדירות מרחבית ומשתנים רבים אחרים אליהם תאי עצב מתכווננים. הארגון המדויק של כל אותם עמודות קליפתיות בתוך ה-V1 נשאר בגדר תעלומה אותה חוקרים כיום.
התאוריה הרווחת בקרב החוקרים כיום היא שתגובות מוקדמות של תאי העצב מורכבות מסדרות רציפות של מסננים בררניים מרחביים-זמניים. בתחום המרחבי, התפקוד של V1 יכול להיות רעיון הדומה למאסף מרחבי, התמרת פורייה מורכבת. באופן תאורטי, יחדיו מסננים אלו יכולים לבצע עיבוד עצבי של תדירות מרחבית, אוריינטציה, תנועה, כיוון, מהירות (כך נמדדת התדירות המרחבית), ועוד הרבה מאפיינים מרחביים-זמניים. ניסויים על תאי עצב ב-V1 מאששים תאוריות אלו, אך גם מעלות שאלות חדשות בנושא.
מאוחר יותר (לאחר 100 מילישניות) תאי העצב ב-V1 רגישים גם לארגון היותר כללי של שדה הראייה. מאפייני תגובות אלו מקורם כנראה מן העיבוד החוזר והנשנה (ההשפעה על אזורי קליפה בעלי רמת עיבוד גבוהה יותר על אזורי קליפה ברמה נמוכה) וההקשרים הצדדיים מן תאי עצב בצורת פירמידה.
המידע החזותי המועבר ל-V1 אינו מקודד מבחינת השימוש בדימויים מרחביים או אופטיים, אלא דווקא כניגוד מקומי. לדוגמה, לתמונה בה צד אחד שחור וחציו השני לבן, קו החלוקה בין השחור והלבן מהווה ניגוד מקומי חזק והוא אינו מקודד, ואילו כמה נוירונים מצפינים מידע בוהק (שחור או לבן כשלעצמו) כאשר המידע מועבר לאזורי הראייה הבאים, הוא מקודד כאותות מופע או תדר הנהפכים ללא מקומיים יותר ויותר. בשלבים מוקדמים אלו של עיבוד המידע הקליפתי, המיקום המרחבי של המידע החזותי נשמר היטב בקרב הקידוד הניגודי המקומי.
שכבות הקליפה

קליפת הראייה העיקרית מחולקת לשש שכבות אופקיות של חומר אפור, הממוספרות מ-1 ועד 6, כאשר לכל אחת מבנה ותפקיד ייחודי (שכבה 1 היא הקרובה ביותר לפני הקליפה ושכבה 6 סמוכה לחומר הלבן שמתחת):
- השכבה הראשונה מורכבת בעיקר מחיבורי אקסונים ודנדריטים.
- שכבות 2 ו-3 מורכבות ממספר רב של תאי עצב השולחים שלוחות אקסונים לאזורי הראייה האחרים. שכבה 2 שולחת פלט ל-V2.
- שכבה מס' 4, אשר מקבלת את רוב הקלט החזותי מן גרעין הברך הצדי, מחולקת גם היא ל-4 שכבות, הנקראות ו-4Cβ, 4Cα, 4B, 4A. שכבה 4B אחראית על התנועה והראייה התלת ממדית, והיא מקבלת קלט מן 4Cα ושולחת פלט ל-V2 ו-V5. שכבת המשנה 4Cα מקבלת את רוב הקלט משכבות תאי העצב המאגנו-תאיים (Magnocellular) בגרעין הברך הצדי, ושולחת פלט לשכבה 4A, בעוד שהשכבה 4Cβ מקבלת את הקלט מן תאי העצב הפארוו-תאיים (Parvocellular), ושולחת פלט לשכבות 2 ו-3, ומעט אל V2.
- שכבות 5 ו-6 מורכבות ממספר רב של תאי עצב השולחים שלוחות אקסונים לעִצבוב גרעין הברך הצדי, ממנו מקבלים קלט. שכבה 5 שולחת פלט לתליל העליון ושכבה 6 בחזרה לגרעין הברך הצדי. יש לציין כי גם בגרעין הברך הצדי שש שכבות.
לכל השכבות מלבד 1 ו-4A יש חיבורים עצביים אל מחוץ לקליפת הראייה העיקרית. שכבות 1 ו-2 מכילות בועות (אזורים עגולים בקליפה המכילות תאי עצב בעלי שדה קלט ניגודי כפול) האחראיות לראייה צבעונית ובכללה צבע וצורה ומקבלות קלט מן שכבה 4Cα ו-4Cβ.
מבנה תאי אזור הראייה הראשוני V1 ותפקידם
לתאים באזור הראייה הראשוני שדה קלט מוארך, ולכן הם מגיבים הכי טוב לגירויים חזותיים ארוכים, כמו קווים. בשנת 1959, צמד החוקרים דייוויד הובל וטורסטן ויזל היו אלה שגילו כי גירוי בצורת קו, גורם לתגובה של תאי ה-V1. ישנו מיתוס האומר כי ויזל והובל גילו תגלית זו בטעות. לפי האגדה, החוקרים הציבו חתול מורדם מול מסך, כאשר עיני החתול פקוחות, ומקרן שקופיות המציג צורות גאומטריות שונות על המסך. הם החדירו כ-45 מיקרו-אלקטרודות לקליפת הראייה, שהיו מחוברות למגבר, כך שכאשר נוצרת תגובה המגבר מעביר צליל סטטי לאוזניות החוקרים, אשר חוזקו תלוי בחוזק התגובה. כאשר הניחו את האלקטרודות ב-V1 התגובות החשמליות שקיבלו מהצורות היו חלשות והבדיקה נערכה זמן מה, כשלפתע נתקעה אחת השקופיות במקרן ונוצר קו זוויתי ממעטפת השקופית, ומיד התקבל רעש חזק באוזניות. לאחר בדיקה ממושכת הבינו החוקרים כי לתאים שונים מתאימות זוויות שונות של הקו. אין שום עדות לכך שסיפור זה הוא נכון, וניתן לראות במאמריהם המסכמים של החוקרים כי מחקריהם התבצעו בצורה מאורגנת ובכוונה תחילה. עם זאת, מרצים באוניברסיטאות ממשיכים לספר סיפור זה מדי פעם כאנקדוטה משעשעת.
צמד החוקרים סיווגו תאים אלו לפי מידת המורכבות של תגובותיהם, המחלק אותם לשתי קבוצות תאים עיקריות: "תא פשוט" ו"תא מורכב".
|
איורים המדגימים שדות קלט בקליפת הראייה העיקרית (בדומה לאיורים המקוריים של החוקרים הובל וויזל):
שדה הקלט של תא פשוט בקליפת הראייה העיקרית. האזור הלבן מייצג אזור מעורר והאפור מעכב. הגירוי (קו שחור) אמנם בגודל ובאורך הנכונים, אך לא באוריינצטיה (כלומר בזווית) הנכונה. לכן, תגובת התא לגירוי (ירי אותות עצביים) תהיה חלשה יחסית.
שדה הקלט של תא מורכב בקליפת הראייה העיקרית. שדה הקלט מורכב כולו מאזור מעורר. הגירוי (קו צר של אור) יעורר תגובה בכל נקודה בשדה הקלט, בכל אורך בו יהיה, כל עוד יהיה באוריינטציה הנכונה. כמו כן, על הגירוי של תא זה לנוע לכיוון החץ למען קבלת תגובה (המהירות בה נע לא משנה את התגובה, כל עוד ניתן לקלוט אותה). כאשר הגירוי אינו באוריינטציה הנכונה, תגובת התא (ירי אותות עצביים) תהיה חלשה או שלא תהיה כלל. |


תאים פשוטים

שדה הקלט בתא פשוט מחולק לשני אזורי משנה; אזור המפעיל מסר מעורר (סינפסה מעוררת, המתחילה את התגובה), הממוקם בחלק הפנימי של התא, ואזור המפעיל מסר מעכב (סינפסה מעכבת, המפסיקה את התגובה) אשר ממוקם בחלקו החיצוני של התא. האזורים המעכבים והמעוררים מסודרים זה לצד זה, ולא במבנה שדה הקלט הרגיל במערכת הראייה, משמע שדה קלט המחולק ל"מרכז" ול"היקף" (צורתם סגלגלה) כאשר פגיעה של גירוי במרכז מעורר תגובה, ובהיקף מעכב.
תאים מסוג זה מגיבים הכי טוב לגירוי באוריינטציה מסוימת (כלומר, תא מסוים יעדיף גירוי הנמצא בזווית מסוימת ה"מתיישרת" עם שדה הקלט שלו), הנמצא יותר באזור המעורר מאשר המעכב, ובעל גודל וכיוון המתאימים לשדה הקלט בתא. מכיוון שתאים שונים מעדיפים אוריינצטיה שונה, יש לכל אוריינטציה נתונה תא פשוט שמגיב לה.
ניתן לראות את העדפת תאים אלו לאוריינטציה מסוימת בעקומה הנקראת "עקומת כיוון האוריינטציה" (orientation tuning curve), הנוצרת על ידי הקלטת תגובות התאים לגירויים בזוויות שונות באמצעות הנחת מיקרו-אלקטרודות על התא.
שדה הקלט בתא פשוט יכול ל"היבנות" בקליפה על ידי תגובות מאורגנות מתאים בגרעין הברך הצדי בעלי שדות קלט הממוקמות לצד פס החוצה את הרשתית.
תאים מורכבים
תאים מורכבים הם התאים השכיחים ביותר באזור הראייה הראשוני, כאשר הם מהווים שלושה רבעים מהתאים בו בערך. גם תאים אלו מגיבים לגירויים באוריינטציה מסוימת אך אינם מקפידים על מיקום הגירויים, כל עוד הם נמצאים בתוך שדה הקלט. תאים מורכבים רבים מגיבים רק כאשר גירוי באוריינטציה התואמת לשדה הקלט נע בכיוון אחד, ולא כאשר הוא נע בכיוון המנוגד.

מחקר נוכחי
מחקרים על קליפת הראייה הראשונית יכולים לכלול הקלטת רמת הדחף העצבי מאלקטרודות הנמצאות בתוך מוחם של בעלי חיים, או על ידי הקלטת אותות אופטיים פנימיים שלהם. כמו כן נמצאים בשימוש החוקרים אמצעים כמו fMRI, לשם הקלטת אותות בתוך ה-V1 של בני אדם או קופי אדם, ומכשיר וידאו המקליט את תזוזת העיניים של הנחקר.
תגלית אחת שפורסמה לאחרונה הנוגעת ל-V1 האנושי היא שאותות שנמדדו על ידי fMRI מראים כמות גדולה מאד של איפיוני קשב[דרוש מקור]. תוצאה זו סותרת מחקר פיזיולוגי על קופי מקוק, המראה שינויים קטנים מאד (אם בכלל) ביריות המקושרות עם איפיוני קשב. מחקרים עם קופי מקוק מתבצעים בדרך כלל על ידי מדידת יריות מניורון בודד. מצד שני, הבסיס העצבי של אותות ה-fMRI הוא קשור ברובו להגברה לאחר הסינפסה (PSP). לכן הבדל זה לא בהכרח מצביע על ההבדל הפיזיולוגי בין קופי מקוק לבני אדם.
מחקר נוסף שמתבצע על V1 שואף לאפיין באופן מלא את תכונותיו הכוונוניים, ולהשתמש בו כאזור מודל למעגל הקליפה הקנוני[דרוש מקור].
פגיעה ב-V1
פגיעה בקליפת הראייה מובילה בדרך כלל למחשך עין (סקוטומה)[3], או חור בשדה הראייה. בצורה מעניינת, מטופלים עם מחשך עין מסוגלים לעיתים קרובות לעשות שימוש במידע חזותי באזור הכתם, על אף אי-יכולתם לקלוט זו ביודעין. התופעה, הנקראת ראייה עיוורת, נחקרת באופן נרחב על ידי מדענים המתעניינים בהתאמה העצבית לתודעה.
נתיבי המידע החזותי מ-V1

קליפת הראייה העיקרית מעבירה את המידע הלאה לאזורי העיבוד המתקדמים בשני נתיבים עיקריים הנקראים הנתיב הגבי והנתיב הגחוני:
- הנתיב הגבי (דורסלי) מתחיל ב-V1, ממשיך דרך אזור הראייה V2, אחר כך לאזור הגבי-התיכון ולאזור הראייה V5 (הידוע גם בשם אזור הראייה MT), ולבסוף לקליפה הקודקודית האחורית (Posterior parietal cortex). הנתיב הגבי, שלעיתים קרוי "נתיב האיפה" ("The Where Pathway"), קשור לתנועה, לתפיסת מיקום עצמים במרחב, ולשליטה על העין והזרועות, במיוחד כאשר נעשה שימוש במידע חזותי על מנת להדריך תנועות מהירות ומתואמות של העיניים (Saccades) או לצורך תנועה כמו הושטת יד[4].
- הנתיב הגחוני (ונטרלי) מתחיל גם הוא ב-V1 ועובר דרך V2, ואחר כך לאזור הראייה V4 ולבסוף מגיע לקליפה הרקתית התחתונה (Inferior temporal cortex). הנתיב הגחוני, שלעיתים קרוי "נתיב המה" ("The What Pathway") קשור לזיהוי צורות ועצמים. כמו כן, הוא קשור באחסון זיכרון לטווח ארוך.
ההפרדה בין הנתיב הגבי לגחוני (הנקראים גם נתיבי המעשה והתפיסה)[4] הוצעה לראשונה על ידי צמד החוקרים אונגרלידר ומישקין (Ungerleider ו-Mishkin)[5] והיא עדיין בגדר עניין הנמצא בוויכוח בקרב החוקרים[דרוש מקור]. הפרדה זו היא ככל הנראה הפשטת יתר של התהליך האמיתי המתרחש בקליפת הראייה.
אזור הראייה השני V2
V2 הוא אזור הראייה השני במרכזיותו בקליפת הראייה. המידע מ-V1 מועבר הלאה ל-V2 בקישורים ישירים ועקיפים, ואזור V2 שולח אותות הלאה לאזורים V3, V4 ו-V5. כמו כן, הוא גם שולח משוב חזרה ל-V1.
V2 מחולק לארבעה רבעים, על פי חלוקה לצד גבי וגחוני ולהמיספרה השמאלית והימנית. יחד, ארבעת החלקים האלה מספקים מפה שלמה של שדה הראייה. תפקידיו של V2 דומים במאפיינים רבים לאלו של V1. גם לו תאים המכווננים על פי מאפיינים פשוטים כמו אוריינטציה, תדירות מרחבית וצבע. תגובותיהם של תאי עצב רבים ב-V2 מווסתות בנוסף על פי מאפיינים מורכבים יותר של הגירוי, כמו מתאר אוריינטציה מתעתעת, והאם הגירוי הספציפי הוא חלק מהתמונה או מהקרקע.
מחקרים שהתבצעו בזמן האחרון מראים כי התאים ב-V2 מראים כמות קטנה של אִפְנוּן קשב ("תשומת לב"), יותר מאשר ב-V1 אך פחות מב-V4. הם מכווננים לדפוסים עם מורכבות בינונית, וכנראה מונעים על ידי אוריינטציות מרובות באזורי משנה שונים בתוך שדה קלט יחיד.
מכלול הראייה השלישי, כולל אזור הראייה השלישי V3
המונח מכלול הראייה השלישי מתייחס לאזור בדיוק לפני V2, הכולל אזור הנקרא בבני אדם אזור V3. המינוח "מכלול" מוצדק מכיוון שקיימת עדיין מחלוקת הנוגעת לגודלו המדויק של אזור V3, כאשר חוקרים מסוימים מציעים כי אזור הקליפה הממוקם לפני ה-V2 נחלק מבחינה תפקודית לשניים או שלושה תתי-אזורים. לדוגמה, החוקר דוד ואן-הסן (David Van Essen) ואחרים (1986) הציעו כי ישנה הבחנה בין "V3 גבי" בחלקן העליון של המיספירות המוח הגדול, לבין "V3 גחוני" (או "האזור הגחוני האחורי" - ventral posterior area) הממוקם בחלקו התחתון של המוח. לאזור V3 הגחוני ו-V3 הגבי יש חיבורים שונים לאזורים אחרים של המוח, וההבחנה ביניהם נראית לעין גם כאשר משתמשים בשיטות שונות של צביעת רקמות.
אזור הראייה הרביעי V4
אזור הראייה החמישי V5 או MT
מקורות
- המאמר: "שדות קלט, אינטראקציות דו-עיניות וארכיטקטורה פונקציונלית בקליפת הראייה בחתולים", מאת הובל וויזל, מתוך כתב העת "The Journal of Physiology", שנת 1962, עמודים 106-154.
- קליפת הראייה העיקרית, מאת מת'יו סצמולסקי (Matthew Schmolesky).
- הספר האלקטרוני: "עיניים, מוח וראייה", מאת דייוויד הובל.
- "קליפת הראייה", מאת פרופסור ג'ורג' מאטר (George Mather), אוניברסיטת סאסקס, אנגליה.
ראו גם
קישורים חיצוניים
הערות שוליים
- ↑ stria of Gennari. הפס נקרא על שמו של פרנצסקו ג'נארי, אנטומיסט איטלקי שחי בין השנים 1750-1795 והיה הראשון לציין אזור זה. מתוך אתר המציג ציוני דרך בניורולוגיה.
- ↑ הגדרה קצרה של קליפת המוח העיקרית, מתוך מילון לפסיכולוגיה.
- ↑ באנגלית Scotoma, זה הוא כתם עיוור בשדה הראייה, מקום בו הראייה חלשה מאד או בלתי קיימת לחלוטין
- ^ 4.0 4.1 סימוכין - המאמר: "Separate pathways for perception and action" (בתרגום חופשי לעברית: "נתיבים שונים לתפיסה ומעשה") מאת Goodale ו-Milner. התפרסם בכתב העת Trends in Neuroscience ב-1992, גיליון ינואר, עמ' 20-25.
- ↑ סימוכין - המאמר: "Analysis of Visual Behavior" (בתרגום חפשי לעברית: "ניתוח התנהגות חזותית") מאת אוגרלידר ומישקין (Ungerleider ו-Mishkin). התפרסם בהוצאת MIT ב-1982. אתר הבית של ההוצאה לאור.
| מערכת הראייה האנושית | ||
|---|---|---|
| העין | רשתית • עדשה • קרנית • קשתית • אישון • תנועות עיניים | |
| מסלולים עצביים | עצב הראייה • תצלובת הראייה • גרעין הברך הצדי • קליפת הראייה • התליל העליון (Superior colliculus) • PPRF • FFA • MLF • FEF | |
| תאים ברשתית | תא קולט אור (קנים, מדוכים) • תא רשתית דו-קוטבי • תא גנגליון • תא אופקי • תא אמקרין | |
| תפיסה חזותית | ראייה • תפיסת עומק • תפיסת צבע • ראיית לילה • שדה ראייה • אשליה אופטית • הכתם העיוור | |
| אביזרים | משקפיים • תרשים לבדיקת ראייה • אוטורפרקטומטר • רטינוסקופ | |

קליפת הראייה25671129