DNA
 | |
| מודל מולקולרי של סליל ה-DNA הכפול. אפור: אטום פחמן, תכלת: אטום חנקן, לבן: אטום מימן, אדום: אטום חמצן, כתום: אטום זרחן. |

DNA (ראשי תיבות באנגלית של Deoxyribonucleic Acid, חומצה דאוקסיריבונוקלאית; בתעתוק לעברית: דנ"א או די־אן־איי) היא מולקולת ענק של חומצת גרעין הבנויה כשרשרת נוקלאוטידים. כל המידע התורשתי הדרוש לבניית החלבונים בתא אצל כל האורגניזמים הידועים, מחיידקים ועד בני אדם, מוצפן במולקולת DNA אחת או יותר, המאורגנות במבנה של סליל כפול, ורצף הנוקליאוטידים בהן ייחודי לכל אורגניזם. גם בחלק מהנגיפים מקודד המידע התורשתי ב-DNA.
אין שני אנשים בעלי אותו ה-DNA. לעיתים תכופות מקובל להשוות את ה-DNA למערכת תוכניות (דוגמת תוכנית מחשב), מכיוון שהוא כולל את כל ההוראות הנחוצות לבניית רכיבי התא, כגון חלבון או מולקולת RNA. כל מקטע משמעותי ב-DNA המכיל מידע לבניית חלבון או הוראות בקרה נקרא "גן". רצף הבסיסים המרכיב את מולקולת ה-DNA מקודד גנים שונים.
כל מולקולת DNA בתאים השונים מורכבת מארבע תת-יחידות של בסיסים חנקניים (נוקלאוטידים), מעין ארבע אבני בניין יסודיות, החוזרות על עצמן לאורכה של המולקולה בצירופים שונים. ארבעת הבסיסים החנקניים הללו הם אדנין (A), גואנין (G), תימין (T) וציטוזין (C), והם מסודרים לאורך הסליל הכפול כך שכל אחד מהם שנמצא בסליל אחד קשור בקשרי מימן עם בן זוג קבוע שנמצא בסליל הנגדי. אדנין (A) וגואנין (G) הם פורינים, ואילו תימין (T) וציטוזין (C) הם פירימידינים. פורין A הוא בן זוג קבוע לפירימידין T, ופורין G הוא בן זוג קבוע לפירימידין C. היחס בין הזוגות הללו בסליל הכפול הוא לפיכך תמיד 1:1, והוא אחד מחוקי צ'רגף. בין אלפים למיליארדים של זוגות בסיסים כאלה מרכיבים את ה-DNA ויוצרים את מבנה הסליל הכפול. בזמן חלוקת התא, הסליל הכפול ארוז בצורה קומפקטית ודחוסה ביותר, במבנה של מארזים הנקראים כרומוזומים.
גילוי מבנה ה-DNA נזקף לזכות המדענים ג'יימס ווטסון האמריקאי ופרנסיס קריק הבריטי בשנת 1953, שזכו בפרס נובל לפיזיולוגיה או לרפואה לשנת 1962 על גילוים זה, יחד עם מוריס וילקינס. בדיעבד התברר שעבודתם התבססה במידה רבה על עבודתה של המדענית רוזלינד פרנקלין, עמיתתו של וילקינס, שנפטרה לפני קבלת הפרס.[1]
מבנה ה-DNA
.jpeg)
מולקולת DNA היא פולימר סלילי הבנוי מיחידה חוזרת. רוחבו של הסליל ואורכה של היחידה החוזרת קבוע. המולקולה היא שרשרת ארוכה, שחוליותיה הם נוקליאוטידים מארבעה טיפוסים, וכל נוקליאוטיד מכיל יחידה של סוכר מסוג דאוקסיריבוז, פוספט (יחידה המכילה זרחן), ובסיס חנקני. ארבעה סוגי בסיסים קובעים את ארבעת טיפוסי הנוקליאוטידים. מספר יחידות אדנין במולקולה זו שווה תמיד לזה של תימין, ומספר יחידות גואנין במולקולה שווה תמיד לזה של ציטוזין.
על פי המודל של ווטסון וקריק, מולקולת DNA היא מבנה דמוי סולם. שני מוטות האורך בסולם בנויים כל אחד כשרשרת של שני מרכיבים המתחלפים ביניהם: סוכר – פוספט – סוכר – פוספט וכן הלאה. אל צידו של כל סוכר קשור בסיס חנקני. כאשר שתי השרשראות ניצבות זו מול זו, פונים הבסיסים החנקניים שבשרשרת האחת לעבר הבסיסים שבשרשרת האחרת. קישור בין בסיסים משתי השרשרות יוצר זוגות של בסיסים המהווים את שלבי הסולם. במצב הטיפוסי שלה, המכונה מבנה מסוג B, המרחק שבין כל מדרגה בסולם הוא 3.4 אנגסטרום והעובי הכולל של מולקולת ה-DNA הוא 20 אנגסטרום (2 ננומטר).
בכל זוג בסיסים שיוצר קשר משתתף בסיס אחד קטן ובסיס אחד גדול. לא כל צירוף של בסיסים אפשרי מבחינה כימית, למעשה מורכבים שלבי הסולם משני סוגים בלבד של צירופי בסיסים חנקניים: אדנין יחד עם תימין, וגואנין יחד עם ציטוזין. אדנין וגואנין השייכים למשפחת הפורינים הם הבסיסים הגדולים, ותימין וציטוזין השייכים למשפחת הפירימידינים הם הבסיסים הקטנים. בין כל שני בסיסים קיימים קשרים כימיים חלשים, קשרי מימן.
בין אדנין ותימין נוצרים שני קשרי מימן ובין גואנין וציטוזין נוצרים שלושה קשרי מימן, ולפיכך הקשר בין גואנין וציטוזין חזק יותר. משמעות הבדל זה היא שזוג של גואנין וציטוזין עמיד יותר בפני שיבושים העלולים להיגרם מהסביבה, כמו מוטציות או התמרה סרטנית. למרות זאת שני הסוגים של זוגות הבסיסים דומים בגודלם, דבר המבטיח שמירה על רוחב סולם קבוע. משמעות החוקיות בזיווג זה של הבסיסים היא בכך שסדר הבסיסים בגדיל אחד של DNA קובע את סדר הבסיסים בגדיל שמולו, המכונה "גדיל משלים". כמו כן, המבנה הדו-גדילי של מולקולת DNA מיוצב על ידי אינטראקציות חלשות בין הבסיסים החנקניים לאורך שרשרת ה-DNA, מסוג "אינטראקציות פאי ארומטיות".
ארגונה של מולקולת DNA במבנה עם השלמה עצמית מהווה את הבסיס לשכפולה מדור לדור, ולהבאת המידע הגנטי הנמצא בה לידי ביטוי. שני גדילי ה-DNA מתפתלים זה סביב זה ויוצרים מולקולה לוליינית המכונה סליל כפול. צימוד משלים זה של שני גדילי DNA מכונה זיווג ווטסון-קריק (ע"ש שני המדענים שתיארו אותה לראשונה), כאשר גדיל אחד מכונה ווטסון והשני מכונה קריק[2].
תפקוד ה-DNA


תהליך ייצור החלבונים על בסיס המידע המוצפן ב-DNA מתרחש בשני שלבים עיקריים: א. שעתוק למולקולות RNA, ב. תרגום לחלבונים.
שעתוק או תעתוק (Transcription)
תהליך השעתוק הוא בניית מולקולת RNA המבוססת על תבנית ה-DNA. ה-DNA עצמו לא מקודד חלבונים באופן ישיר. במקום זאת הוא מקודד את מולקולות ה-RNA שבאמצעותן מיוצרים כל החלבונים הנחוצים לתפקוד התא. ה-RNA הוא מולקולה חד-גדילית הדומה במבנה שלה למבנה גדיל יחיד של מולקולת DNA, והנבדלת ממנה בכך שיחידות הסוכר בנוקלאוטידים שלה הן ריבוז במקום דיאוקסיריבוז (חוסר באטום חמצן אחד), וכן בכך שרנ"א מכיל אוראציל, ו-DNA מכיל תימין.
התחלת התהליך היא כאשר קטע מתאים בסליל הכפול של ה-DNA נפתח, ואחד הגדילים שבו משמש כתבנית לסינתוז ה-RNA. האנזים האחראי על התהליך נקרא RNA פולימראז (RNA polymerase). אנזים זה נקשר לאזור מיוחד ב-DNA הנקרא קָדָם (פרומוטר, promoter).
שלב השעתוק הוא מהחשובים בתא, משום שבאמצעות השליטה בו נקבע אילו חלבונים ייווצרו בתא באותו זמן. באמצעות שליטה מבוקרת בהפעלת השעתוק של קטעי DNA שונים, כל אחד בעיתו, נוצרת השונות בין התאים השונים, ונקבעים השלבים השונים בחיי התא. לדוגמה, באמצעות בקרה מתאימה ייווצר החלבון שנותן את הצבע לעיניים רק בתאים של הקשתית, בעוד שבתאים שאינם תאי קשתית מושתק השעתוק של קטע ה-DNA המכיל את הגן לצבע העיניים.
תרגום (Translation)
בשלב התרגום נעשית בניית החלבון על-פי ה-RNA. כל שלשה של בסיסים ברצף ה-RNA (ולפניו ב-DNA) מתורגמת לחומצה אמינית אחת ברצף החלבון. רצף הבסיסים הראשוני ב-DNA קובע, על כן, את רצף חומצות האמינו בחלבון, ועקב כך את מבנהו ותפקודו.
מיקום וסידור
באורגניזמים פרוקריוטיים חסרי גרעין נמצא ה-DNA בציטופלזמה, שהיא הנוזל התוך תאי, והוא מעגלי ורציף. רוב החומר התורשתי ביצורים אלו שוכן על גבי מולקולת DNA טבעתית אחת.
באורגניזמים אוקריוטיים, שתאיהם בעלי גרעין, ה-DNA מסודר בגרעין התא, שם הוא נמצא בצורת כרומטין, שהוא סליל DNA מלופף סביב חלבונים הנקראים היסטונים. חלבונים אלה מאפשרים לו להיות ארוז בצורה יעילה ומקטינים את הנפח שהוא תופס. בדיקת מיקום וסידור הכרומטין על רצף הDNA, כלומר גילוי האזורים שהם הטרוכרומטין ואאוכרומטין, נעשית בין היתר על ידי DNA footprinting.
גילויים חשובים בחקר ה-DNA
פענוח המבנה
 ערך מורחב – גילוי מבנה ה-DNA
ערך מורחב – גילוי מבנה ה-DNA
עד לתחילת שנות החמישים, ידעו מדענים על קיומם של גנים, המעבירים תכונות תורשתיות, אך סברו כי החלבונים, שהם מולקולות מורכבות ביותר, הם אלה הנושאים את המידע התורשתי. ה-DNA היה נחשב לחומר פשוט ומשעמם המשמש כמרכיב מבני של הכרומוזומים וזאת מפני שטעו לחשוב שארבע האותיות של ה-DNA מופיעות בו בסדר מחזורי קבוע. הבקטריולוג א.ת. איוורי ממכון רוקפלר בניו יורק ערך ניסוי חשוב שהראה כי אפשר להעביר תכונות בין חיידקים באמצעות העברת DNA. ניסויו של איוורי הצביעו על כך שהגנים מצויים ב-DNA.
התפנית הגדולה בהבנת מבנה ה-DNA התרחשה כאשר רוזלינד פרנקלין הצליחה ליצור הדמיה דו-ממדית של סליל ה-DNA על ידי קריסטלוגרפיה בקרני רנטגן. התמונה הטובה ביותר מכונה "תצלום 51". ווטסון וקריק השיגו את התצלום בלא ידיעתה, בעזרתו של מוריס וילקינס, עמיתהּ של פרנקלין בקינגס קולג' והצליחו בעזרתו לפענח את המבנה התלת-ממדי של מולקולת ה-DNA. הם הבינו שמדובר במבנה סליל דו-גדילי, חישבו את מספר הבסיסים בכל סיבוב של הסליל ובנו דגם תלת ממדי של ה-DNA מלוחות מתכת.
פענוח המבנה הפריך את ההנחה בדבר סדר מחזורי של בסיסים במולקולת ה-DNA. האותיות יכולות להופיע בכל סדר שהוא בתוך הסליל הכפול, בלי שהמבנה עצמו יושפע כלל (כל עוד עיקרון זיווג האותיות של שתי השרשרות נשמר). בעקבות זאת, ניתן היה מיד להציע כי סדר האותיות בכל מולקולת DNA הוא למעשה מכיל את המידע התורשתי. גדולתו של דגם המבנה של ה-DNA הדו-גדילי שהוצג על ידי ווטסון וקריק היא בכך שמבנה מולקולה זה מתאים למילוי תפקידה כמולקולת התורשה: המבנה מציע הסבר למנגנון השכפול העצמי של המולקולה, לאופן שבו המידע הגנטי אצור בה ואף לדרך שבה שינויים במידע מועברים בתורשה.
פענוח הקוד הגנטי
בשנת 1961 ערכו פרנסיס קריק וסידני ברנר סדרת ניסויים ובהם הפעילו על DNA ממקור נגיפי חומר הגורם לסילוק זוגות בסיסים מה-DNA ועקב כך לשינויים תורשתיים. סדרת ניסויים זו, הובילה להבנת תהליך התרגום. אובדן זוג בסיסים, או שני זוגות בסיסים מהרצף, הוא למעשה החסרת חלק ממילה בקידוד ה-DNA, ובנוסף לזה גורם להזזת מסגרת הקריאה של שלשות הבסיסים בדי-אן-איי. רצף הבסיסים משנה עקב כך את משמעותו ומתורגם לרצף חלבון שונה. לעומת זאת, אובדן של שלושה זוגות בסיסים גורם להחסרת חומצת אמינו אחת, אך יתר החלבון נשאר תקין.
בתוך חמש שנים נתגלתה המשמעות של כל 64 הצרופים האפשריים לרצף של שלושה זוגות בסיסים: 61 מצירופי השלשות השונים מקודדים ל-20 חומצות האמינו השונות המרכיבות את החלבונים בתא (מספר צירופים שונים עשויים לקודד לאותה חומצת אמינו), ובנוסף שלוש שלשות שאינן מקודדות לחומצת אמינו אלא משמשות כאות סיום. צירופים אלו, של שלשות בסיסי חומצות גרעין, מכונים קודונים. תגליות מאוחרות יותר הראו שכאשר יש רצף ארוך שמשותף לשני גנים, אחד הגנים יהיה בחפיפה לשני ואז יקטע וההמשך שלו יהיה אחרי אזור שלא משתתף בקידוד. במצב כזה הגן הראשון יקודד ברציפות וכאשר מתבצע שימוש בקידוד של הגן המקוטע, רצף הדי אן איי יתפתל ללולאה שמסתירה את המקטע שלא צריך להשתתף ביצירת החלבון. כיום יודעים שהקוד הזה אוניברסלי לכל היצורים החיים ולנגיפים[3].
ראו גם
- PNA
- תיקון DNA
- דנ"א אוריגמי
- ריצוף DNA
- חידת ערך C
- שכפול עצמי
- זיווג בסיסים
- רצף נשנה
- בדיקת סמני DNA
- פרופיל DNA
- DNA footprinting
- נזקי DNA בעקבות קרינת UV וקרינת רנטגן
לקריאה נוספת
- ג'יימס ווטסון, הסליל הכפול, ידיעות ספרים, 2003
- עדי מרקוזה-הס, ביולוגיה עכשיו, ספרי תל אביב
קישורים חיצוניים
| מיזמי קרן ויקימדיה |
|---|
- ארז גרטי, ה-DNA - חוברת ההפעלה של הגוף, במדור "מאגר המדע" באתר של מכון דוידסון לחינוך מדעי, 30 ביולי 2009
- צבי ינאי, על גנים ועל נגיפים, מחשבות 53, דצמבר 1984, עמ' 10–19
 אסף רונאל, פיצוח האתגר הבא בחקר הדנ"א צובר תאוצה. ההשלכות יהיו מרחיקות לכת, באתר הארץ, 13 במרץ 2019
אסף רונאל, פיצוח האתגר הבא בחקר הדנ"א צובר תאוצה. ההשלכות יהיו מרחיקות לכת, באתר הארץ, 13 במרץ 2019- DNA, באתר אנציקלופדיה בריטניקה (באנגלית)
הערות שוליים
- ^ שמעון שוקן, הסליל הכפול: איך שני פרחחים לקחו את כל הקופה, 23.3.2005, באתר רשימות; במקור, פורסם ב-2004 במדור ספרים של הארץ
- ^ Cartwright, Reed; Graur, Dan (8 פבר' 2011). "The multiple personalities of Watson and Crick strands". Biology Direct. 6: 7. doi:10.1186/1745-6150-6-7. PMC 3055211. PMID 21303550.
{{cite journal}}: (עזרה)תחזוקה - ציטוט: date and year (link) - ^ למיטוכונדריה ולכלורופלסט יש קוד גנטי שונה אולם הם אינם יצורים עצמאיים אלא אברונים. כמו כן ישנם חיידקים שכאשר יש בחלל התא שלהם סלניום מתיונין, אחד מקודוני הסיום שלהם הופך לקודון עבורה.
| חומצות גרעין | ||
|---|---|---|
| אבני בניין | נוקלאוזיד • נוקלאוטיד • דהאוקסינוקלאוטיד | 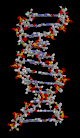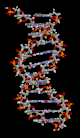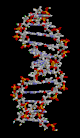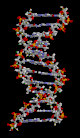
|
| פורין | אדנין (A) • גואנין (G) | |
| פירימידינים | תימין (T) • ציטוזין (C) • אורציל (U) | |
| RNA | רנ"א מקודד: mRNA • pre-mRNA
רנ"א שאינו מקודד תרגום: tRNA • rRNA • tmRNA בקרה: miRNA • siRNA • piRNA • RNAi עריכת רנ"א: snRNA • snoRNA | |
| DNA | cDNA | |

33960632DNA